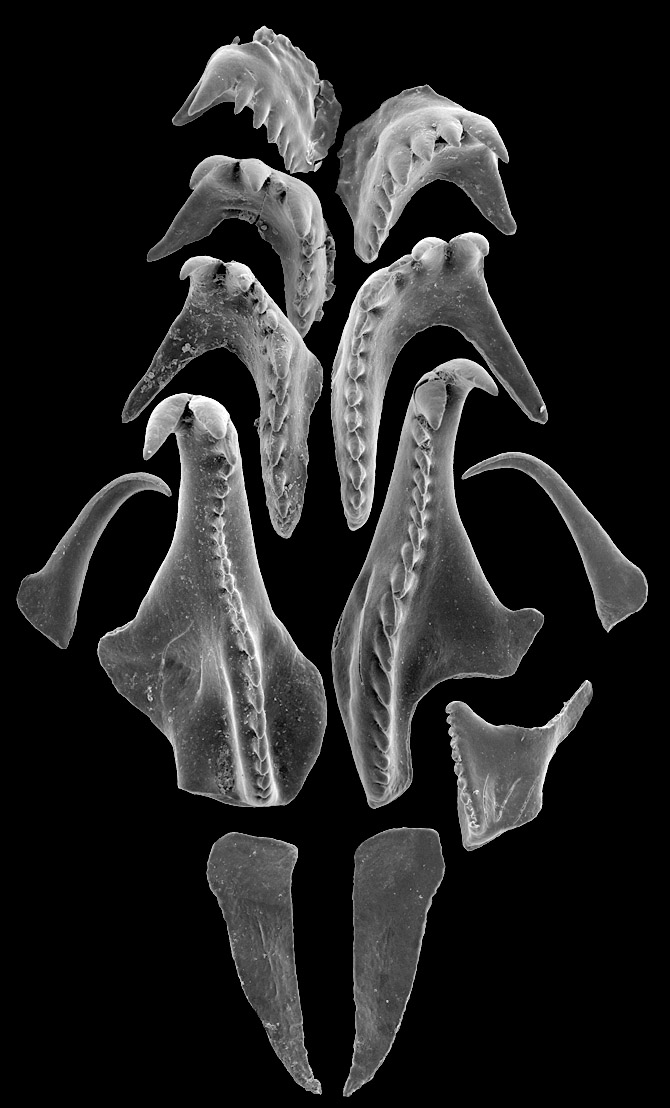
Ordoviitsiumi ajastu lõuguslase Ramphoprion rekonstrueeritud lõua-aparaat, mis koosneb vähemalt 12st osisest ehk skolekodondist. Pildistatud skaneeriva elektronmikroskoobiga, Olle Hintsi kollektsioonist (TTÜ Geoloogia Instituut). Pildi kõrgus u 2.5 mm.
Skolekodondid — lühidalt
- Nimi: skolekodondid; ingl k scolecodonts
- Süstemaatiline kuuluvus: skolekodondid on hulkharjasusside (Annelida, Polychaeta) lõuaaparaadi elemendid (“hambad”)
- Kuju: erineva kujuga üksikud hambad ja hammastega plaadikesed
- Koostis: kollakas- või tumepruun kuni must orgaaniline aines
- Suurus: mikrofossiilid, enamasti 200–1000 μm, harva kuni 5 mm
- Stratigraafiline levik: Ülem-Kambrium – tänapäev
- Eluviis: mereline, bentos, vastsed planktilised
- Kogumine ja uurimine: lahustusmeetod (kõik happed), stereomikroskoopia, mikroskoopia, SEM
- Tähtsus: hulkharjasusside evolutsiooni selgitamine, keskkonnaindikaatorid, vähem biostratigraafias
- Eestis: laialt levinud Ordoviitsiumi-Siluri kivimites, hea säilivus, suhteliselt hästi uuritud
Sissejuhatus
Skolekodondid kujutavad endast hulkharjasusside ehk polüheetide (Annelida, Polychaeta) lõua-aparaadi mikroskoopilisi elemente, mis koosnevad vastupidavast orgaanilisest ainesest ja kuuluvad alates Ordoviitsiumist tavalisemate happeresistentsete mikrofossiilide hulka.
Polüheetide lõua-aparaate on võimalik kasutada perekondade ja liikide eristamiseks. Kuna pehmekehaliste polüheetide tervikfossiilid (body fossils) on üliharuldased, siis kujutavad skolekodondid endast peamist tõendit selle tänapäevalgi laialt levinud ja ökoloogiliselt olulise loomarühma arengust ja rollist geoloogilises minevikus. Eesti aluspõhja karbonaatkivimites on skolekodondid ühed sagedamini kohatavad mikrofossiilid (kitiinikute, konodontide, akritarhide ja ostrakoodide kõrval).
Morfoloogia ja terminoloogia
Kõik üksikuna leitud skolekodondid pärinevad lõua-aparaatidest, kuhu kuulub mitmeid erineva kuju ja suurusega elemente. Lõua-aparaatide ehituses eristatakse viit põhitüüpi: plakognaatne, labidognaatne, prionognaatne, ktenognaatne, ja ksenognaatne aparaat. Valdav enamus taksoneid omab kolme esmamainitud tüüpi aparaate. Plakognaatse ja ktenognaatse aparaaditüübi eristamine on mõneti problemaatiline. Samuti esineb üleminekulise tüübiga aparaate labidognaatse ja prionognaatse tüübi vahel.

Skolekodontide morfoloogia ja terminid.
Klassifikatsioon
Tänapäevaste polüheetide klassifikatsioon tugineb valdavalt pehme keha morfoloogial ja molekulaarsetel tunnustel, mis fossiilsena ei säilu. Detailsed tööd fossiilse materjali põhjal on näidanud, et lõuaaparaadid ja skolekodondid on samuti kasutatavad perekondade ja liikide eristamiseks. Siiski ei ole täpselt teada kas lõuaaparaatide alusel püstitatud fossiilsed liigid vastaksid täpselt tänapäeval kasutatavale liigikontseptsioonile.
Fossiilsete skolekodontide kaasaegne klassifikatsioon tugineb sugukonna ja perekonna tasemel lõua-aparaadi ehitusele, erinevate elementide arvule ja morfoloogiale aparaadis. Liikide püstitamiseks ja eristamiseks vajalikud tunnused esinevad ka üksikutel aparaadi elementidel. Samas ei ole kaugeltki mitte kõik aparaadi elemendid võrdse taksonoomilise väärtusega – väiksed elemendid ja üksikud hambakesed ei ole enamasti liigini, aga sageli ka perekonnani määratavad. Kõige diagnostilisemad elemendid on üldjuhul tagamaksillid (MI), 2. maksillid (MII), vahel ka basaalplaadid.
Aparaadi üldise ehituse järgi on eristatud 5 aparaaditüüpi, mis kujutavad endast mitteformaalset taksonoomilist taset: labidognaadid, plakognaadid, prionognaadid, ktenognaadid ja ksenognaadid. Iga aparaaditüübi hulka võib kuuluda mitmeid sugukondi, mida eristab üksteisest elementide erinev arv, paigutus, või märkimisväärselt erinev kuju ja funktsioon.
Stratigraafiline levik
Vanimad skolekodondid on teada Kanada Ülem-Kambriumi kihtidest. Üksikuid leide, sh Eestist, on kindlaks tehtud Alam-Ordoviitsiumi Tremadoci lademest. Skolekodontide arvukus ja mitmekesisus kasvas märgatavalt Kesk-Ordoviitsiumi alguses; siis ilmusid ka esimesed keerukama aparaaditüübiga vormid (labidognaadid ja prionognaadid), mis valdavad kuni tänapäevani. Lõugsete polüheetide mitmekesistumist on seni kõige põhjalikumalt analüüsitud Ordoviitiumi ajastul (Hints & Eriksson, 2008). Ordoviitsiumi ja Siluri polüheedifaunad olid üsna sarnased, Devonis ilmus aga rida uusi sugukondi. Teada on ka, et Permi lõpus (Permi-Triiase massiline väljasuremine) kadus enamik Paleosoikumile iseloomulikke vorme. Mesosoikumi kivimites on skolekodondid vähem arvukad, kuid ka vähem uuritud. Võimalik, et kaasaegsetele sarnased vormid omasid väiksema fossiliseerumispotentsiaaliga lõua-aparaate. Ökoloogia Tänapäevaste polüheetide ökoloogia on väga mitmekesine, kuid enamikele vormidele on iseloomulik bentiline eluviis (välja arvatud vastsena ning vahel ka sigimisperioodil). Polüheedid on eranditule merelised loomad, neid võib kohata väga erinevates keskkondades alates litoraalist kuni abüssaalini. Polüheetide toitumisviisid on samuti mitmekesised ulatudes sette- või hõljumitoidulistest vormidest kuni raipesööjate ja kiskjateni.
Uurimismetoodika
Skolekodondid kuuluvad happeresistentsete fossiilide hulka ja nende kogumisel on valdavaks lahustusmeetodi kasutamine. Skolekodontide orgaanilisest ainest kest talub hästi kõiki happeid ning seetõttu saab neid eraldada ka terrigeensetest kivimitest kasutades fluorhapet.
Skolekodontide eraldamiseks lahustusjäägist kasutatakse märgsõelumist (sõela Ø 45-125 μm, valdavalt 63 μm). Edasise metoodika osas kasutatakse nii märg- kui kuivnoppimist. Märgnoppimine ja skolekodontide püsisäilitamine glütseriinipreparaadis sobib paremini väikeste eksemplaride ja tervikuna säilunud lõuaaparaatide uurimiseks. Seda meetodit kasutas nn Poola koolkond ning see leiab rakendamist ka Eestis. Rootsi uurijad eelistavad seevastu kuivnoppimist ja säilitamist kaamerates sarnaselt konodontide puhul kasutatava metoodikaga. Selle viisi eeliseks on võimalus üksikeksemplarid sorteerida ja liimiga püsivalt kaamerapõhjale fikseerida. Viimane on aeganõudev tegevus kuid lihtsustab oluliselt edaspidist uurimistööd (näiteks muutlikkuse uurimist).
Skolekodontide sorteerimisel ja määramisel kasutatakse stereomikroskoopi (suurendused 20–100×). Pildistamiseks ja mikrostruktuuride uurimiseks on vajalik skaneeriva elektronmikroskoobi (SEM) rakendamine.
Tähtsus ja praktiline kasutatavus
Paljude hulkharjasusside evolutsioon oli suhteliselt aeglane, mõne perekonna eluiga võib ulatuda üle 200 milj aasta (selliseid taksoneid tuntakse kui "elavaid fossiile"). Siiski on skolekodontide hulgas ka suhteliselt lühikese stratigraafilise levikuga liike, kelle kasutatavust biostratigraafias pärsib uurituse ebapiisav tase. Siiski, bentilise eluviisi tõttu omavad skolekodondid stratigraafilist potentsiaali eelkõige neis kihtides, kus paremad indeksfossiilid puuduvad.
Skolekodondid Eestis
Eesti Ordoviitsiumi ja Siluri kivimites on skolekodondid üheks sagedamini kohatavateks mikrofossiilideks. Eesti (ja Baltika paleokontinendi) seni vanimad skolekodondid pärinevad Pakerordi lademe graptoliitargilliidist (Türisalu kihistu) ning noorimad Ülem-Siluri Ohesaare lademest. Kambriumist ja Devonist ei ole Eestis seni skolekodonte leitud. Ordoviitsiumi-Siluri kihtidest on Eestist teada u 200 eri liiki, millest osa on veel formaalselt kirjeldamata. Eesti skolekodontide kollektsioon on maailmas üks suuremaid. Kõige tavalisemad skolekodondid Eestis kuuluvad perekondadesse Oenonites, Mochtyella, Pistoprion, Kettnerites, Pteropelta, Ramphoprion, Atraktoprion jt.